Kommentar til boka 'Evolusjon eller Kristen tro' -ja takk begge deler
Kap2. Spor i våre gener etter evolusjon -og evt. endring i arvematerialet
(I stykket under benyttes forkortelsen NDT for Neo-Darwinistisk Teori.)
Påstand i boka: "Ja, vår felles utviklingshistorie støttes også av fossiler i oss alle, mer presist i våre gener."
Kommentar: Antallet mulige pseudogener har i løpet av få år, sunket fra 95% til under 20%, og synker fortsatt. Så hvorfor feiler evolusjonære forutsigelser så ofte i forbindelse med pseudogener? Grunnen er at evolusjonære forutsigelser ser genomet som et produkt av tilfeldig prøvende og feilende prosesser. Det mest produktive synet på et genom i et forskningsperspektiv, ville være å forvente vedvarende funksjonalitet og utrolig bio-engineering, som produkt fra en allmektig og vis Skaper.
"Ett eks. er ett gen kalt L-gulano- γ-lakton-oksidase (kort 'GULO'-genet), som gjør at pattedyr kan danne c-vitaminer i kroppen, bortsett fra mennesker og andre primater, som må få c-vitaminet via mat."
Mange av de pattedyrene som ikke er i stand til å danne askorbin syre (vitamin C), har regioner i genomet sitt, som antas å korrespondere med deler av det funksjonelle GULO-genet, som er funnet i de pattedyrene som er i stand til å danne GULO, og således vitamin C.
Bilde 1. GULO pseudogen
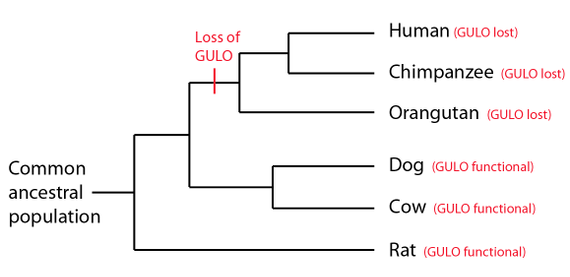 Evolusjonister har promotert GULO-pseudogenet som bevis for delte feil mellom sjimpanse og menneske, i det de har benyttet et lite antall av høyst pre-selekterte data. Deres bevisførsel er knyttet til enkelte innsettinger i ett ekson, knyttet til et fungerende GULO-gen i rotte, ikke i relasjon til en hel sekvens av GULO-gener. Vi skal se under at dette ikke 'holder vann'. I motsetning til evolusjonære påstander om ødeleggelser av vitamin C -og dets nettverk, så er de artsmessig begrenset i sine genomiske signaturer (1). De viser kjennetegn på å være uavhengige begivenheter. I en nyere studie av forfatteren til dette stoffet, så viser genomiske data at multiple tap av GULO-pseudogener i menneske, sjimpanse, og gorilla opptrådte uavhengig i hver art. Disse er assosiert med degenererte regioner, som inneholder en vid variasjon av ombyttbare gjentagende sekvenser (1). Ved reduksjonsdeling (meiosis) kan det skje uforutsette ting, som f.eks. å gjøre om sekvens-sletting i gener. GULO gen-regionen synes å ha blitt påvirket av dette. Kombinert med det faktum at mutasjoner i dette (vitamin-C) genet, ikke ville være dødelig, vil et mutert gen bli tolerert og sendt videre til etterkommerne. I denne sammenheng er det et poeng at ombyttbare elementer i det området der GULO-gen eksoner gikk tapt, er unike for mennesker, sjimpanse og gorilla, noe som indikerer at begivenhetene skjedde uavhengig av hverandre.
Evolusjonister har promotert GULO-pseudogenet som bevis for delte feil mellom sjimpanse og menneske, i det de har benyttet et lite antall av høyst pre-selekterte data. Deres bevisførsel er knyttet til enkelte innsettinger i ett ekson, knyttet til et fungerende GULO-gen i rotte, ikke i relasjon til en hel sekvens av GULO-gener. Vi skal se under at dette ikke 'holder vann'. I motsetning til evolusjonære påstander om ødeleggelser av vitamin C -og dets nettverk, så er de artsmessig begrenset i sine genomiske signaturer (1). De viser kjennetegn på å være uavhengige begivenheter. I en nyere studie av forfatteren til dette stoffet, så viser genomiske data at multiple tap av GULO-pseudogener i menneske, sjimpanse, og gorilla opptrådte uavhengig i hver art. Disse er assosiert med degenererte regioner, som inneholder en vid variasjon av ombyttbare gjentagende sekvenser (1). Ved reduksjonsdeling (meiosis) kan det skje uforutsette ting, som f.eks. å gjøre om sekvens-sletting i gener. GULO gen-regionen synes å ha blitt påvirket av dette. Kombinert med det faktum at mutasjoner i dette (vitamin-C) genet, ikke ville være dødelig, vil et mutert gen bli tolerert og sendt videre til etterkommerne. I denne sammenheng er det et poeng at ombyttbare elementer i det området der GULO-gen eksoner gikk tapt, er unike for mennesker, sjimpanse og gorilla, noe som indikerer at begivenhetene skjedde uavhengig av hverandre.
Dette voldte at forfatterne i et nylig evolusjonær forskningsprosjekt, fullstendig oppgav de å bruke store segmenter i GULO-gen regionen til artsmessig forskning (2). De benyttet i stedet segmenter som ga data passende for standard slutninger til evolusjonsmodellen. Altså det samme oppsettet med pre-selekterte data, brukt i så mange andre studier (cherry-picking).
Sammenlignet med sjimpanse og gorilla, er de 28.800 basene i menneskelig GULO pseudogen-region bare h.h.vis. 84% og 87% like (1). Og de 13.000 basene med DNA som går forut for levninger av menneskelig GULO-gen, -knyttet til tap av to hoved-eksoner, er bare 67% og 73% like h.h.vis. sjimpanse og gorilla. Disse resultatene ligger utenfor intervallet til felles avstamnings paradigmet og gorillaen er betydelig mer lik menneske i denne regionen, enn sjimpanse er, noe som ytterligere motbeviser den vanlig oppfattede artsmessig avstamningen.
På grunn av små endringer i ekson-regionen til menneskelig GULO-gen, det er få eller ingen, så har begivenheten antagelig nylig funnet sted. Om det skjedde for millioner av år siden, ville eksonene vært mye mer nedbrutt. I tillegg indikerer nåværende ENCODE-data i menneskelig GULO-region at det epigenetiske maskineriet til genomet har gjenkjent og tagget det som ødelagt og har lukket det ned fra å bli transkribert, for å beskytte tilhørende regioner som inneholder aktive gener (1).
GULO pseudogenet er er klart kjennetegn på genetisk diskontinuitet mellom mennesker og aper, ikke på arv av felles mistak. Artsmessig bestemte gen-degraderings-begivenheter dukker opp som et felles tema, assosiert med genetisk entropi og diskontinuitet mellom artene, ikke med makro-evolusjon.
Påstand i boka: "Blant slike spor av evolusjonstrinn, som ikke lenger har sin opprinnelige funksjon, er haler hos mennesker"
Kommentar: Idéen om at alle menneskelige embryoer har en "hale" er falsk hvis det som er ment, har noe å gjøre med en stamfar med hale. Det kommer fra det faktum at i løpet av fjerde eller femte uke av den normale prosessen med utvikling, utvikler alle mennesker aposterior-forlengelse av fosteret. Da utvikles muskel-skjelettstruktur utover endetarmsåpning, noe som hjelper å utfolde kroppens plan og nervesystemet. Dette er imidlertid ikke på grunn av noe tilbakedannet 'hale-DNA'; Det er en kritisk fase av programmert menneskelig embryonal utvikling, i det ryggstreng og nevrale rør strekker seg over det meste av denne halelignende strukturen.
 Denne struktur, bare meget overfladisk lik en hale, fungerer som en slags mal eller stillas, som framkaller eller styrer dannelsen av andre strukturer ved presise forløp i den senere utvikling (for eksempel virker ryggstreng som en mal for celler som utvikler seg til ryggvirvler). Når disse systemene er ferdige, sikrer genetisk programmering fjerning av den opprinnelige strukturen, som ikke lenger er nødvendig. Og dette er ikke unikt for nettopp denne posteriore forlengelse; mange slike strukturer dannes og reabsorberes under normal menneskelig utvikling. 'Annals of Anatomy' beskriver det på denne måten (de kaller det dessverre 'hale'): "Under normal menneskelig utvikling dannes en rekke forbigående strukturer og går deretter helt tilbake. En av de mest fremtredende strukturer som regress under utviklingen er den menneskelige hale ... Til å begynne med er den menneskelige halen sammensatt av hale-spire fostervev som differensierer til .. sekundære nervetråder, ryggstreng og endetarm."(22)
Denne struktur, bare meget overfladisk lik en hale, fungerer som en slags mal eller stillas, som framkaller eller styrer dannelsen av andre strukturer ved presise forløp i den senere utvikling (for eksempel virker ryggstreng som en mal for celler som utvikler seg til ryggvirvler). Når disse systemene er ferdige, sikrer genetisk programmering fjerning av den opprinnelige strukturen, som ikke lenger er nødvendig. Og dette er ikke unikt for nettopp denne posteriore forlengelse; mange slike strukturer dannes og reabsorberes under normal menneskelig utvikling. 'Annals of Anatomy' beskriver det på denne måten (de kaller det dessverre 'hale'): "Under normal menneskelig utvikling dannes en rekke forbigående strukturer og går deretter helt tilbake. En av de mest fremtredende strukturer som regress under utviklingen er den menneskelige hale ... Til å begynne med er den menneskelige halen sammensatt av hale-spire fostervev som differensierer til .. sekundære nervetråder, ryggstreng og endetarm."(22)
Bilde. Aposterior-forlengelse av fosteret-hjelp til utfolding av nervesystemet
En analogi ville være som en steinmur utgjør en buegang. Fordi steinene i en bue ikke vil stå av seg selv til sluttstenen er plassert for å fullføre det, bygger murere en "bukk" (et tre-stillas i form av en bue) under den, slik at den vil støtte steinene i det buen bygges. Ved ferdigstillelse fjernes 'bukken', og den ferdige buegangen er selvbærende. Med mindre du hadde sett buen bygges, hadde du kanskje aldri visst at stillaset var der. Og det er et forbilde på omtanke og design, ikke noen 'på måfå sammen-kastet opphopning av leftovers'.
Fullt fungerende haler?
Om en analyser erklæringen om at mennesker ofte er født med "fullt fungerende" haler, kan det best oppnås ved å definere hva en ekte hale er, hvilke funksjoner haler har, og den strukturen som kreves for denne funksjonen, og så sammenligne dem med såkalte 'menneskelig haler'. Husk at argumentet om 'velfungerende' ikke er om mennesker noen gang har blitt født med en utvekst hvor en hale kan være (hvis vi hadde vært utformet med haler). Heller ikke selv om en utvekst skyldes utviklingsmessig unormal forløp, inneholder muskler (som hvis koblet til nerver vil trekke seg sammen, og dermed føre til at utstikket flytter seg). Det er om mennesker noensinne faktisk har haler sammenlignet med dyrehaler, eller ikke (-enten i embryo-form eller når de er født).
Dyrehaler er fullt utviklet strukturelle ryggvirvler, som fortsetter forbi de bakre hoftene med hensiktsmessig festede muskler, nerver som aktiverer disse musklene med aktuelle nervebaner for kontroll av halen (-hele veien til, og i hjernen), og andre nødvendige myke vev som er tilstede i anatomisk passende forhold til alle disse.
Bilde. Struktur i ryggmargsbrokk
Til tross for de inntrykk man får, har ingen såkalte 'virkelige haler' (husk, disse har muskler) i mennesker noensinne blitt funnet å inneholde bein eller brusk (23,24), og selv om 'pseudo-haler' er blitt funnet å inneholde bein, har de ikke inneholdt virvler (Coyne uttalelse om at noen menneskelige haler har virvler, ser ut til å være falsk, fra alle kilder vi har vært i stand til å vurdere). Så alle disse ur-vedheng ligner ikke i det hele tatt noen reell dyrehale strukturelt. Og ingen mennesker er født med en slik abnormitet er i stand til å bruke sitt hale-vedheng for noen av funksjonene ovenfor. Dermed blir påstanden om at mennesker ofte er født med 'fullt fungerende' haler, villedende og falsk.
Lege og kirurg Dr. Michael Egnor som har faktiske kirurgisk erfaring med denne type tilstand, uttalte følgende om 'menneske haler':
Ingen av dem -og ingen av rapporter i litteraturen som jeg vet om, er virkelige haler.-Michael Egnor (nestleder ved Institutt for Nevrologisk kirurgi, og direktør, Pediatric Nevrokirurgi ved State University i New York ved Stony Brook). "Ingen av dem-og ingen rapporter i litteraturen som jeg vet om, er faktiske haler. En hale har ryggvirvler, er en fortsettelse av halebenet, har utviklet muskler, nerver og andre myke vev, etc. Vedheng som er beskrevet i litteraturen, og alle de vedheng som jeg har operert, er misdannede (mesenchymale) vev, ..(spes. faguttrykk).., som bærer en overfladisk likhet med en 'hale'. Ingen har strukturen av en hale, selv i rudimentær form, og ingen av de jeg har operert ble festet til halebenet på den måten en hale er." (25)
Se mer om dette her.
Kap2. Spor i våre gener etter - endring i arvematerialet
Påstand i boka (s.66): "Når et område i DNA..med én eller flere gener kopieres dobbelt, kan det muteres uavhengig av originalsekvensen."

Kommentar: Evolusjonister tror at de fleste gener oppstår ut fra pre-eksisterende foreldre-gener, via gjentatte dupliserings-begivenheter over lange tidsperioder. Etter at de er duplisert, tenkes det at de omstokkes og rearrangeres til å forme nye, relaterte gener. Denne idéen bygger på at mange gener, selv innen samme organisme, deler like gen-regioner. Gener som deler signifikante likhetsnivåer med hverandre, sies å representere gen-familier.
Det er rett nok at gen-duplisering kan opptre ved anledning. Det er et antall av seriøse problem med denne modellen som en farbar, evolusjonær drivkraft. For det første innebærer det å endre rekkefølgen på de fleste gener, fysiske sykdommer og utviklings avvik som leder til enten skader eller dreper organismen. For det andre, er antatt dupliserte gener små integrerte stykker av et høyst komplekst og inter-avhengig gen-nettverk. Å tenke seg at å kopiere ett av disse komplekse stykkene, og så tilfeldig skyfle disse rundt omkring, for å skape et nytt og fantastisk integrert gen, er absurd i de fleste tilfeller. Programmerere vet at om de gjør det med programkode, blir det mange feilmeldinger..
Bilde 2. Hypotese ved gen-duplisering
Det er nå velkjent blant molekylære biologer at gener fungerer i integrerte, komplekse, overlappende, interavhengige og utsøkt kontrollerte nettverk. Videre er disse gen-nettverkene dynamiske og svarer til all slags stikkord og signaler, som konstant kontrolleres av celle-systemet. Det finnes ikke motstykke til dette kompleksitets- og effektivitets-nivået i systemer som er planlagt og designet mennesker. Biologer kan ta lærdom av gode programmerere. Gode programmerere vil gjenbruke deler av sin kode og syntaks, i ulike deler av sitt arbeid og også mellom ulike software prosjekter. At duplisering av ulike sekvenser dukker opp i ulike gener, tilsvarer et standard prinsipp i programmering.
Utsagn i boka: "Det hevdes at man har for stor tiltro til mutasjoner og genetiske mekanismer. Forskningen har stagnert, og man bør snarest komme seg ut av en blindgate."
Kommentar: Det hjelper ikke at 'en rekke biologer jobber med modeller basert på kunnskap fra fagfelt som atferdsbiologi og fysiologi, økologi og kunstig intelligens', om alt skal sees gjennom evolusjonsteoriens linser, som har en tendens til å gi et vridd bilde av virkeligheten. Det virker for øvrig ikke som den norske debatten er særlig heftig, når eneste nevnte eks. hevder: 'Evolusjonsbiologien-trenger-ingen-revolusjon'.
Se mer: her.
Videre påstand i boka: "Det førstnevnte skjer når et noen områder i et DNA kan bli overført til andre ved noe som kan sammenlignes med h.h.vis cut&paste og copy&paste. Det førstnevnte skjer når et område i DNA blir fjernet og flyttet til et annet sted (transponering). Den andre når et område blir kopiert og flyttet (replikativ transponering). Her er det flere viktige poenger. Ett er at denne type endringer kan spores i arvematerialet, slik at det er mulig å se hvilke arter som har en felles forhistorie."
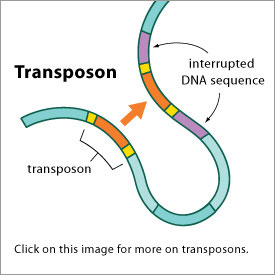 Kommentar: Allan Rogers (2011) skrev at transposoner er brukt til å estimere fylogeni, og at de vitner om sannhet ved felles avstamning. Han argumenterer at' siden transposoner ikke er funksjonelle og setter seg selv tilfeldig inn i genomet, så kan de bli brukt til å etablere et fylogenetisk tre'. Han skriver (s27): 'det er ekstremt usannsynlig at to transposoner noen sinne vil sette seg inn på samme sted. Det innebærer at to individer som deler samme transposon også må ha samme stamfar.'
Kommentar: Allan Rogers (2011) skrev at transposoner er brukt til å estimere fylogeni, og at de vitner om sannhet ved felles avstamning. Han argumenterer at' siden transposoner ikke er funksjonelle og setter seg selv tilfeldig inn i genomet, så kan de bli brukt til å etablere et fylogenetisk tre'. Han skriver (s27): 'det er ekstremt usannsynlig at to transposoner noen sinne vil sette seg inn på samme sted. Det innebærer at to individer som deler samme transposon også må ha samme stamfar.'
Bilde 3. Transposoner -funksjonelle og på foretrukne steder
Det viser seg imidlertid at dette premisset er usant:
1) Transposoner er funksjonelle (Emera og Wagner, 2012). Vi har tidligere sett at de kan være ansvarlige for å reorganisere genomet, som et tilpasset svar på input fra omgivelsene.
2) De setter seg ikke tilfeldig inn. J. Shaphiro som har studert transposoner i mer enn 35 år, rapporterer at transposoner setter seg inn på foretrukne steder (Shaphiro, 2011). Det samme har andre eksperter på transposoner gjort (Levy et al. 2010). Som en konsekvens er ikke to transposoner på samme sted i genomet, noen indikasjon på at de deler en stamfar. Så mye for Roger's forsøk på å kreve bevis for felles avstamning fra transposoner.
Påstand i boka: "Ikke minst vises det i forbindelse med fenotypisk plastisitet"
Kommentar: Gitt fenomenet fenotypisk plastisitet, at organismer med samme genotype (genetisk konstitusjon/arveanlegg) har ulike fenotyper. Ut fra det kan en spørre hvor mye av variasjonen i organismer er resultat av 'preprogrammering' i motsetning til tilfeldige mutasjoner? Om gjentatte variasjoner stammer fra preprogrammering, hvor befinner den nødvendige informasjonen seg og hvordan kommer den til uttrykk?
 Krabber lever av snegler, men kan ikke spise snegler med tykt skall. Snegler kan på ett eller annet vis vite om krabber er i området. I nærvær av krabber får de tykkere skall (Stearns 1989). Denne tilpasningen hjelper klart å beskytte snegler fra krabber. Snegler er selv rovdyr som lever av et dyr i småkrepsgruppen (rankefoting-barnacle). Når disse senser snegler, beskytter de seg ved å vokse inn i en krumbøyd stilling som hindrer sneglene å spise dem. Når det ikke finnes snegler omkring, går rankefotingen tilbake til sin opprinnelige form (Stearns 1986). Det forfatteren (Dr Lee Spetner) foreslår her, er at organismer har en innebygd evne til å tilpasse seg omgivelsene. Han foreslår at i den grad evolusjon foregår, foregår det på individnivå hos organismen. Dette forslaget skiller seg skarpt fra tesen til NDT, som holder på at evolusjon inntreffer bare på populasjonsnivå. Organismer inneholder informasjonen som setter dem i stand til å utvikle en fenotype tilpasset en variasjon av omgivelser. Tilpasningen kan inntreffe ved en endring i genomet gjennom en genetisk endring trigget av omgivelsene, eller det kan inntreffe uten noen genetisk endring.
Krabber lever av snegler, men kan ikke spise snegler med tykt skall. Snegler kan på ett eller annet vis vite om krabber er i området. I nærvær av krabber får de tykkere skall (Stearns 1989). Denne tilpasningen hjelper klart å beskytte snegler fra krabber. Snegler er selv rovdyr som lever av et dyr i småkrepsgruppen (rankefoting-barnacle). Når disse senser snegler, beskytter de seg ved å vokse inn i en krumbøyd stilling som hindrer sneglene å spise dem. Når det ikke finnes snegler omkring, går rankefotingen tilbake til sin opprinnelige form (Stearns 1986). Det forfatteren (Dr Lee Spetner) foreslår her, er at organismer har en innebygd evne til å tilpasse seg omgivelsene. Han foreslår at i den grad evolusjon foregår, foregår det på individnivå hos organismen. Dette forslaget skiller seg skarpt fra tesen til NDT, som holder på at evolusjon inntreffer bare på populasjonsnivå. Organismer inneholder informasjonen som setter dem i stand til å utvikle en fenotype tilpasset en variasjon av omgivelser. Tilpasningen kan inntreffe ved en endring i genomet gjennom en genetisk endring trigget av omgivelsene, eller det kan inntreffe uten noen genetisk endring.
Bilde 4. Snegler kan tilpasse seg omgivelsene
Noen spørsmål går på effektiviteten til evolusjonære mekanismer. Kan disse mekanismene forklare ulike design og kanskje også en designende intelligens? Hvor store endringer kan ikke-styrte prosesser som mutasjoner og utvalg danne? Kan de produsere nye proteiner, for ikke å si nye anatomiske? Eller er det kanskje andre design-mønstre kjent fra programvare-design eller mekanisk bygging som kan forklare trekk vi finner? Motsatt om ikke-styrte evolusjonære mekanismer er tilstrekkelige for å gjøre rede for opphavet til alle nye livsformer: er det mulig at gjennomtrengende tegn på design i høyere livsformer var preprogrammert for å folde seg ut fra livets opprinnelse av?
Utsagn i boka: "Muligens kan noe av kritikken, også fra ID-hold, stimulere til å tenke annerledes på enkelt områder".
Kommentar: Ja, kanskje i forbindelse med: C-verdi paradokset
Forskning på En-Ikke-Tilfeldig-Evolusjonshypotese (EITEH) kan føre til løsning på det som kalles C-verdi paradokset. Massen av DNA i en organismes (haploide) kjønnscelle, kalles organismens "C-verdi". C-verdien er vanligvis uttrykt i picogram (pg), som er en billionte-del av et gram. Det er 978 millioner base-par i et picogram. C-verdien til mennesker er ca 3.5 pg, som tilsvarer ca 3.4 milliarder basepar. DNA i cellene til en organisme, blir vanligvis forstått å inneholde informasjon nødvendig for dens utvikling og funksjon. Det synes derfor fornuftig å anta at mengden til en organismes DNA, er et slags mål på dens kompleksitet. Men det synes ikke å være et konsistent forhold mellom C-verdien og den tilsynelatende kompleksiteten til organismen. Denne inkonsistensen har vært betraktet som et paradoks. Det syntes problematisk at C-verdien for noen dyr var betydelig større enn for mennesket.Verdien for den blå-flekkede salamanderen (Ambystoma laterale), er 81 pg, eller 23 ganger større enn for mennesket. C-verdi for amerikansk hulesalamander (Necturus lewisi) er 120 pg, eller 34 ganger større enn for mennesket. Det er ingen grunn til å anta at utvikling og virkemåte til disse dyrene er mer komplekse enn for et menneske. Det synes dermed merkelig at disse dyrene skulle ha så mye mer DNA enn ett menneske. Denne gåten er kjent som C-verdi paradokset (Thomas, 1971).

 Like merkelig er det at selv noen nære slektninger til dyr eller planter har svært annerledes C-verdier. F.eks. de to paddene (Scaphiopus couchii) og den europeiske (Bombina bombina). Den første har en C-verdi på nær 1, mens den andre har en C-verdi på mer enn 12. Likedan for de to froskene, den utsmykkede hule-frosken (Ceratophrys ornata), den første har en C-verdi på omkring 1, mens en horn-utsmykket slektning (Limnodynastes ornatus) har C-verdi på mer enn 13. Det er mange andre eks. på parallelle arter med lignende morfologi, som har vidt ulike C-verdier. Variasjonen i C-verdi over hele klassen amfibier har en faktor på 120. (-fra 'Animal Genome Database: genomesize.com) Hvorfor skulle veldig like dyrearter, som er meget like, ha slike ulike C-verdier? Om én av dem kan utvikle seg og fungere med mindre DNA-innehold, hvorfor trenger den andre gjøre det?
Like merkelig er det at selv noen nære slektninger til dyr eller planter har svært annerledes C-verdier. F.eks. de to paddene (Scaphiopus couchii) og den europeiske (Bombina bombina). Den første har en C-verdi på nær 1, mens den andre har en C-verdi på mer enn 12. Likedan for de to froskene, den utsmykkede hule-frosken (Ceratophrys ornata), den første har en C-verdi på omkring 1, mens en horn-utsmykket slektning (Limnodynastes ornatus) har C-verdi på mer enn 13. Det er mange andre eks. på parallelle arter med lignende morfologi, som har vidt ulike C-verdier. Variasjonen i C-verdi over hele klassen amfibier har en faktor på 120. (-fra 'Animal Genome Database: genomesize.com) Hvorfor skulle veldig like dyrearter, som er meget like, ha slike ulike C-verdier? Om én av dem kan utvikle seg og fungere med mindre DNA-innehold, hvorfor trenger den andre gjøre det?
Bilde 5. To frosker med kvotient 13 i C-verdi
'Konsensus-svaret' ligger i det formodede uberegnelig resultat av tilfeldige mutasjoner og naturlig seleksjon. Svaret ligger i begrepet junk-DNA (Eddy, 2012, Gregory, 2005). 'Konvensjonell visdom' er at ulikhet i C-verdier kan tilskrives store mengder av ubrukelig eller 'søppel-DNA' i noen arter og mindre i andre.. Men disse svarene kommer til kort, i forhold til ENCODE-programmet, som rapporterte at 'søppel-DNA' ikke er søppel. Darwinisters argument er sirkulært: eksistensen av 'søppel-DNA' sies å løse C-verdi paradokset, mens C-verdi paradokset blir brukt som et argument for validiteten av 'søppel-DNA'. Men et sirkelargument er ingen løsning. For darwinister må C-verdi problemet være et paradoks.
 Imidlertid synes det som EITEH kan løse C-verdi paradokset. Den innebygde kapasiteten til å svare på omgivelses-input, krever informasjon, og denne informasjonen ville mest sannsynlig ligge i genomet. Forfatteren vil kalle DNA-et som bærer denne informasjonen for I-DNA, der I står for Ikke-tilfeldig. Vanligvis ville ikke I-DNA være aktivt. Om en skulle slette en del av det, ville en vanligvis ikke se noen effekt. Vi nevnte tidligere en ultra-bevart del av DNA som ble fjernet fra ei mus, uten synlig virkning (Ahituv et al. 2007) At delen var ultra-bevart, tydet på en viktig effekt {-etter som den ikke ble fjernet via naturlig-seleksjon-oversetters kommentar}. Siden fjerning av DNA-et ikke ga synlig utslag, må det henge sammen med noe annet enn normalt liv og utvikling. Effekten ville bli merkbar kun i nærvær av en egnet stimulus fra omgivelsene, som bare kunne inntreffe i endrede omgivelser. Forfatteren foreslår at I-DNA er ekstra DNA i arter med høy C-verdi. Arter som må tilpasse seg til en vid variasjon av omgivelses-forhold, burde ha et større volum av DNA. De som bare tilpasser seg til forhold innenfor et smalt spekter, skulle ha mindre I-DNA. Det er eks. på et interessant forskningsprosjekt knyttet til EITEH.
Imidlertid synes det som EITEH kan løse C-verdi paradokset. Den innebygde kapasiteten til å svare på omgivelses-input, krever informasjon, og denne informasjonen ville mest sannsynlig ligge i genomet. Forfatteren vil kalle DNA-et som bærer denne informasjonen for I-DNA, der I står for Ikke-tilfeldig. Vanligvis ville ikke I-DNA være aktivt. Om en skulle slette en del av det, ville en vanligvis ikke se noen effekt. Vi nevnte tidligere en ultra-bevart del av DNA som ble fjernet fra ei mus, uten synlig virkning (Ahituv et al. 2007) At delen var ultra-bevart, tydet på en viktig effekt {-etter som den ikke ble fjernet via naturlig-seleksjon-oversetters kommentar}. Siden fjerning av DNA-et ikke ga synlig utslag, må det henge sammen med noe annet enn normalt liv og utvikling. Effekten ville bli merkbar kun i nærvær av en egnet stimulus fra omgivelsene, som bare kunne inntreffe i endrede omgivelser. Forfatteren foreslår at I-DNA er ekstra DNA i arter med høy C-verdi. Arter som må tilpasse seg til en vid variasjon av omgivelses-forhold, burde ha et større volum av DNA. De som bare tilpasser seg til forhold innenfor et smalt spekter, skulle ha mindre I-DNA. Det er eks. på et interessant forskningsprosjekt knyttet til EITEH.
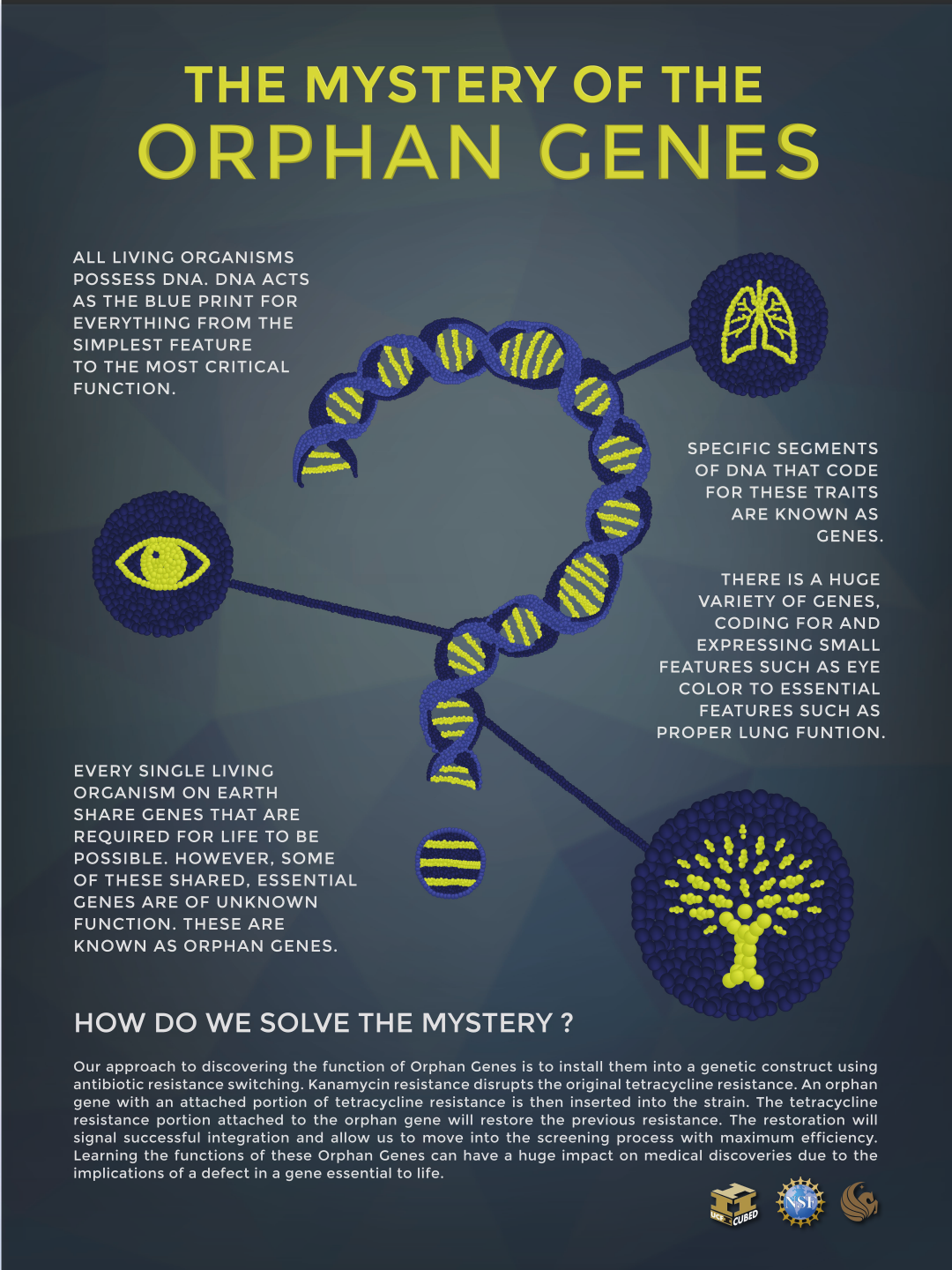
Bilde 6. Orfan-gener-plutselig dukker de opp-hvorfra?
Påstand i boka: "Naturvitere flest, 'uansett livssyn', mener at naturlig evolusjon fra et felles utgangspunkt, er den uten sammenlikning beste forklaringen på livets utvikling og mangfold."
Kommentar: Grunnlaget for at 'den uten sammenligning beste forklaringen på livets utvikling og mangfold .. evolusjonsteorien', har problemer med å forklare flere forhold, f.eks. plustelige, raske forandringer (her), kanskje etterfulgt av lange perioder med stillstand (stasis-paradokset), mange parallelle, uavhengige utviklingsforløp ('konvergent evolusjon'), samt de såkalte 'orfan'-genene. Samt problemene med naturlig seleksjon knyttet til å bevare ikke-funksjonelle endringer, og at intet nytt tilføres i forbindelse med naturlig seleksjon. Denne luker bare vekk ikke hensiktsmessige mutasjoner/variasjoner.
Orfan-gener
Takket være moderne genetikk, har forskere klart å avdekke en forbløffende kategori gener: 'taksonomisk begrensede gener' (orfan-gener). Disse genene er identifisert i mennesket, og knyttet til funksjoner over hele kroppen. Det spesielle ved dem, er at de plutselig dukker opp, uten noen form for forløpere i aper eller noe annet dyr. De passer ikke det hypotetisk evolusjonære paradigmet. De er ofte knyttet til miljømessig tilpasning. Når forskere analyserer tilsynelatende dupliserte gener hos mennesker og aper, så er det i de fleste tilfeller utstrakte forskjeller mellom antall kopier, gen-lokasjoner og sekvenser genomene i mellom(3-4).
Mislykkede forsøk på forklaring: De Novo gen-syntese
 Det spesielle ved orfan-gener er basert på det fakutm at de dukke plutselig opp (uten forløpere) og ikke har lignende motparter i andre genom. Evolusjonister benekter begrepet konstruert design i biologiske systemer. Derfor må de finne en begrunnelse for hvordan slike smart designede DNA-sekvenser tilfeldig ble dannet i meget nær evolusjonær tid..
Det spesielle ved orfan-gener er basert på det fakutm at de dukke plutselig opp (uten forløpere) og ikke har lignende motparter i andre genom. Evolusjonister benekter begrepet konstruert design i biologiske systemer. Derfor må de finne en begrunnelse for hvordan slike smart designede DNA-sekvenser tilfeldig ble dannet i meget nær evolusjonær tid..
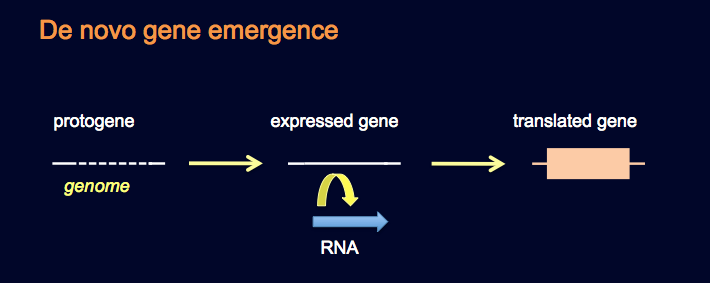
Bilde 7. 'Det er lov å forsøke': de-novo gen-syntese
Det som foreløbig er dukket opp, er en forestilling som kalles 'de novo gene synthesis' ('gen syntese på ny'). Det skal visstnok skje ved tilfeldig, mutasjonsdannelser av fullt fungerende gener av ikke-kodende DNA, som skjer meget hurtig. Siden 'evolusjon må være sant, så må de ha oppstått fra ikke-kodende DNA, for hvor ellers skulle de stamme fra.' {evolusjon i hullene - i henhold til oversetters mening.} Selv om det er skrevet ulike forskningsartikler som tilsynelatende beviser 'gen syntese på ny', så er de i virkeligheten eksempler på en sirkulær slutning (tautologi): 'gen syntese på ny' må være sant, siden orfan-gener eksisterer, og grunnen til orfan-gener må være 'gen syntese på ny'.
Bilde 8. Menneskelig genom -i seg selv et bevis

Referanser:
1. Tomkins, J.P. 2013. The human GULO pseudogen- evidence for Evolutionary Discontinuity and Genetic Entropy. Answers Research Journal.
2. Lachapelle MY, Drouin G. 2011. Inactivation dates of the human and guinea pig vitamin C genes. Genetics 139(2): 199-207
3. Dennis, M.Y. et al, 2012. Evoution of human specific neural SGGAP2 genes by incomplete segmental duplication. Cell 149:912-922
4. Charrier C. et al. 2012. Inhibition of SRGAP2 function by its human-specific paralogs induces neotony during spine maturation. Cell 149:923-935
Utvalgt og redigert av Asbjørn E. Lund